生物化學與分子生物學/ATP的生成、儲存和利用
| 醫學電子書 >> 《生物化學與分子生物學》 >> 生物氧化 >> ATP的生成、儲存和利用 |
| 生物化學與分子生物學 |
|
|
ATP幾乎是生物組織細胞能夠直接利用的唯一能源,在糖、脂類及蛋白質等物質氧化分解中釋放出的能量,相當大的一部分能使ADP磷酸化成為ATP,從而把能量保存在ATP分子內。
ATP為一游離核苷酸,由腺嘌呤、核糖與三分子磷酸構成,磷酸與磷酸間借磷酸酐鍵相連,當這種高能磷酸化合物水解時(磷酸酐鍵斷裂)自由能變化(G)為30.5KJ/mol,而一般的磷酸酯水解時(磷酸酯鍵斷裂)自由能的變化只有8至12KJ/mol,因此曾稱此磷酸酐鍵為高能磷酸鍵,但實際上這樣的名稱是不夠確切的,因為一種化合物水解時釋放自由能的多少取決于該化合物整個分子的結構,以及反應的作用物自由能與產物自由能的差異,而不是由哪個特殊化學鍵的破壞所致,但為了敘述及解釋問題方便,高能磷酸鍵的概念至今仍被生物化學界采用。
ATP是一高能磷酸化合物,當ATP水解時首先將其分子的一部分,如磷酸(Pi)或腺苷酸(AMP)轉移給作用物,或與催化反應的酶形成共價結合的中間產物,以提高作用物或酶的自由能,最終被轉移的AMP或Pi將被取代而放出,ATP多以這種通過磷酸基團等轉移的方式,而非單獨水解的方式,參加酶促反應提供能量,用以驅動需要加入自由能的吸能反應,ATP水解反應的總結如下:
ATP——→ADP+Pi
或ATP——→AMP+PPi
(焦磷酸)
目錄 |
一、ATP的生成方式
體內ATP生成有兩種方式
(一)底物水平磷酸化(substrate level phosphorylation)
底物分子中的能量直接以高能鍵形式轉移給ADP生成ATP,這個過程稱為底物水平磷酸化,這一磷酸化過程在胞漿和線粒體中進行,包括有:

(二)氧化磷酸化(oxidative phosphorylation)
氧化和磷酸化是兩個不同的概念。氧化是底物脫氫或失電子的過程,而磷酸化是指ADP與Pi合成ATP的過程。在結構完整的線粒體中氧化與磷酸化這兩個過程是緊密地偶聯在一起的,即氧化釋放的能量用于ATP合成,這個過程就是氧化磷酸化,氧化是磷酸化的基礎,而磷酸化是氧化的結果。
機體代謝過程中能量的主要來源是線粒體,既有氧化磷酸化,也有底物水平磷酸化,以前者為主要來源。胞液中底物水平磷酸化也能獲得部分能量,實際上這是酵解過程的能量來源。對于酵解組織、紅細胞和組織相對缺氧時的能量來源是十分重要的。
二、氧化磷酸化偶聯部位的測定
確定氧化磷酸化偶聯部位通常用兩種方法。
(一)P/0值測定
P/0值指在氧化磷酸化過程中消耗一克原子氧所消耗的無機磷的克原子數,或者說消耗一克原子氧所生成的ATP的克分子數。P/0值實質上指的是呼吸過程中磷酸化的效率。
測定P/0值的方法通常是在一密閉的容器中加入氧化的底物、ADP、Pi、氧飽和的緩沖液,再加入線粒體制劑時就會有氧化磷酸化進行。反應終了時測定O2消耗量(可用氧電極法)和Pi消耗量(或ATP生成量)就可以計算出P/0值了。在反應系統中加入不同的底物,可測得各自的P/0值,結合我們所了解的呼吸鏈的傳遞順序,就可以分析出大致的偶聯部位了。
表6-3 離體線粒體的P/0比值
| 底物 | 呼吸的組成 | P/0比值 | 生成ATP數 |
| (1)β-羥丁酸 | NAD+→FMN→CoQ→Cyt→O2 | 2.4-2.8 | 3 |
| (2)琥珀酸 | FAN→CoQ→Cyt→O2 | 1.7 | 2 |
| (3)抗壞血酸 | Cyt→Cytaa3→O2 | 0.88 | 1 |
| (4)細胞色素c | Cytaa3→O2 | 0.61-0.68 | 1 |
從上表可以看出P/0值為小數,由于線粒體的偶聯作用在離體條件下不能完全發揮,故可認為實際的ATP生成數是他們所接近的正整數值。
比較表中的(1)和(2),呼吸鏈傳遞的差異是在CoQ之間,兩者ATP的生成數相差1,所以這個ATP的生成部位一定在NAD→CoQ之間。
比較表中(2)和(3),呼吸鏈傳遞的差異是在Cyt c之間,兩者ATP的生成數相差1,所以這個ATP的生成部位在CoQ→Cyt c之間。
比較表中(3)和(4),生成的ATP數均為1,呼吸鏈傳遞的區別是在Cyt c→Cyt aa3,故Cytc→ aa3不存在偶聯部位,而在Cyt aa3→O2之間存在著一個偶聯部位。
(二)根據氧化還原電位計算電子傳遞釋放的能量是否能滿足ATP合成的需要
氧化還原反應中釋放的自由能△G'O與反應底物和產物標準氧化還原電位差值(△E'O)之間存在下述關系:△G'O=nF△E'O
式中n為氧化還原反應中電子轉移數目,F為法拉弟常數(23.062千卡/克分子.伏特,或96500庫侖/克分子)。
一克分子ATP水解生成ADP與Pi所釋放的能量為7.3千卡,凡氧化過程中釋放的能量大于7.3千,均有可能生成一克分子ATP,就是說可能存在有一個偶聯部位,根據上式計算,當n=2時,△E'O=0.1583V時可釋放7.3千卡能量,所以反應底物與生成物的標準氧化還原電位的變化大于0.1583V的部位均可能存在著一個偶聯部位。
從圖6-11可以看出,在NAD→CoQ,Cyt b→Cyt c和Cyt aa3→O2處可能存在著偶聯部位。必須明確,這種計算的基礎是反應處在熱力學平衡狀態,溫度為25℃,pH為7.0,反應底物和產物的濃度均為1克分子,這種條件在體內是不存在的。因此這一計算結果只能供參考。
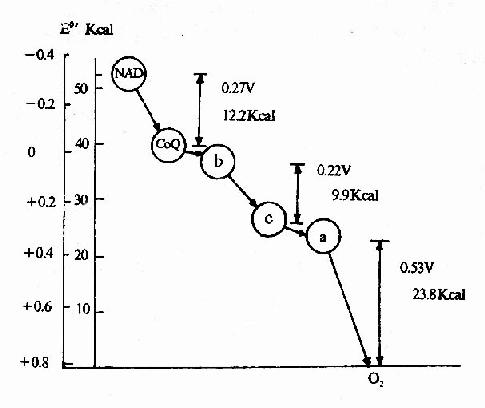
圖6-11 呼吸鏈中電子對傳遞時自由能的變化
綜上所述,呼吸鏈中電子傳遞和磷酸化的偶聯部位可用圖6-12表示。
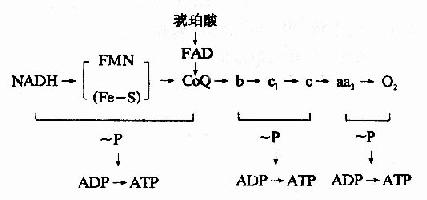
圖6-12 偶聯部位示意圖
呼吸鏈磷酸化的全過程可用下述方程式表示
NADH+H++3ADP+3Pi+1/2O2→NAD++3ATP+4H2O
FADH2+2ADP+2Pi+1/2O2→FAD+2ATP+3H2O
三、氧化磷酸化中ATP生成的結構基礎
ATP是由位于線粒體內膜上的ATP合成酶催化ADP與Pi合成的。ATP合成酶是一個大的膜蛋白質復合體,分子量在480?00kD,是由兩個主要組成(或稱因子)構成,一是疏水的F0,另一是親水的F1,又稱F0F1復合體。在電子顯微鏡下觀察線粒體時,可見到線粒體內膜基質側有許多球狀顆粒突起,這就是ATP合成酶,其中球狀的頭與莖是F1部分,分子量為350?/P>380kD,由α3、β3、γ、δ、ε等9種多肽亞基組成,β與α亞基上有ATP結合部位;γ亞基被認為具有控制質子通過的閘門作用;δ亞基是F1與膜相連所必需,其中中心部分為質子通路;ε亞基是酶的調節部分,F0是由3?個大小不一的亞基組成,其中有一個亞基稱為寡霉素敏感蛋白質(oligomycinsensitivityconferringprotein,OSCP),此外尚有一個蛋白質部分為分子量28kD的因子,F0主要構成質子通道(見圖6-13)。
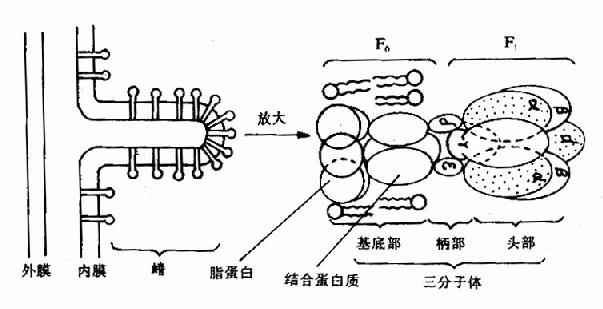
圖6-13 線粒體內膜上三聯體(三分子體)結構示意圖
四、氧化磷酸化的偶聯機制
有關氧化磷酸化的偶聯機理已經作了許多研究,目前氧化磷酸化的偶聯機理還不完全清楚,50年代Slater及Lehninger提出了化學偶聯學說,1964年Boear又提出了構象變化偶聯學說,這兩種學說的實驗依據不多,支持這兩種觀點的人已經不多了。目前多數人支持化學滲透學說(chemiosmotic hypothesis),這是英國生化學家P.Mitchell于1961年提出的,當時沒有引起人們的重視,1966年他根據逐步積累的實驗證據和生物膜研究的進展,逐步地完善了這一學說。
氧化磷酸化的化學滲透學說的基本觀點是:
1.線粒體的內膜中電子傳遞與線粒體釋放H+是偶聯的,即呼吸鏈在傳遞電子過程中釋放出來的能量不斷地將線粒體基質內的H+逆濃度梯度泵出線粒體內膜,這一過程的分子機理還不十分清楚(見圖6-14)。
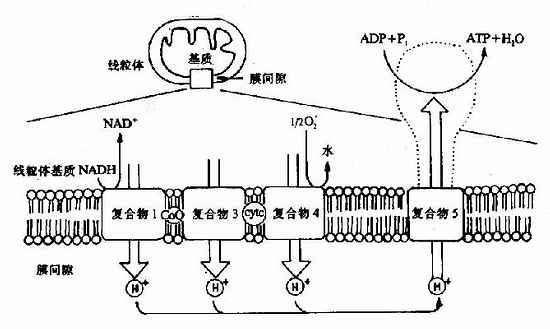
圖6-14 電子傳遞與質子傳遞偶聯(注:復合物Ⅱ未顯示)
2.H+不能自由透過線粒體內膜,結果使得線粒體內膜外側H+濃度增高,基質內H+濃度降低,在線粒體內膜兩側形成一個質子跨膜梯度,線粒體內膜外側帶正電荷,內膜內側帶負電荷,這就是跨膜電位△ψ。由于線粒體內膜兩側H+濃度不同,內膜兩側還有一個pH梯度△pH,膜外側pH較基質pH約低1.0單位,底物氧化過程中釋放的自由能就儲存于△ψ和△pH中,若以△P表示總的質子移動力,那么三者的關系可用下式表示:
△P=△ψ-59△pH
3.線粒體外的H+可以通過線粒體內膜上的三分子體順著H+濃度梯度進入線粒體基質中,這相當于一個特異的質子通道,H+順濃度梯度方向運動所釋放的自由能用于ATP的合成,寡霉素能與OSCP結合,特異阻斷這個H+通道,從而抑制ATP合成。有關ATP合成的分子機制目前還不十分清楚。
4.解偶聯劑的作用是促進H+被動擴散通過線粒體內膜,即增強線粒體內膜對H+的通透性,解偶聯劑能消除線粒體內膜兩側的質子梯度,所以不能再合成ATP。
總之,化學滲透學說認為在氧化與磷酸化之間起偶聯作用的因素是H+的跨膜梯度。
每對H+通過三分子體回到線粒體基質中可以生成一分子ATP。以NADH+H+作底物,其電子沿呼吸鏈傳遞在線粒體內膜中形成三個回路,所以生成3分子ATP。以FADH2為底物,其電子沿琥珀酸氧化呼吸鏈傳遞在線粒體內膜中形成兩個回路,所以生成兩個ATP分子。
自從Mitchell提出化學通透學說以來,已為大量的實驗結果驗證,為該學說提供了實驗依據。
美國Cohen等人于1978年使用完整的大鼠肝細胞作實驗材料,以核磁共振(nuclearmagneticresonance,NMR)的方法直接觀察到完整細胞中胞液與線粒體基質之間存在H+跨膜梯度,胞液的pH值比線粒體基質的pH值低0.3單位,用解偶聯劑處理,或用氮氣代替氧氣切斷氧的供應,那么胞液和線粒體基質之間的pH梯度消失。
嗜鹽菌(halobacteriumhaloblum)是一種能在高濃度鹽溶液中生長的細菌,該菌中有一種結合蛋白質,稱為菌紫質(bacteriorhodopsin),菌紫質能將光能轉換成化學能。有人使用嗜鹽菌作實驗,在無O2的情況下用光照射嗜鹽菌,盡管無氧化作用,菌體內仍維持一定的ATP濃度,若加入解偶聯劑或加入磷酸化抑制劑DCC,則菌體內ATP濃度降低;而加入呼吸抑制劑抑制電子傳遞,即不影響ATP合成,ATP濃度不變,這說明電子傳遞和H+運動是可以分開加以研究的,嗜鹽菌為研究化學滲透學說的H+運動提供了一個理想的模型。于是,有人分離嗜鹽菌的菌紫質,并將其重組在人工脂質體中,然后用光照射,可測得跨膜電位為120mV(內負外正),同時膜外側H+濃度增高,膜內外△pH約為1.8單位,可以算出總的質子移動力約為△P=-120mV-59×1.8mV=226mV,若再將牛心線粒體內膜重組在此脂質體中,光照后可使ADP+Pi生成ATP,這說明質子跨膜梯度可以經過線粒體內膜的三分子體將H+跨膜梯度中儲存的能量轉變為ATP分子中的化學能。
五、氧化磷酸化抑制劑
氧化磷酸化抑制劑可分為三類,即呼吸抑制劑、磷酸化抑制劑和解偶聯劑。
(一)呼吸抑制劑 這類抑制劑抑制呼吸鏈的電子傳遞,也就是抑制氧化,氧化是磷酸化的基礎,抑制了氧化也就抑制了磷酸化。呼吸鏈某一特定部位被抑制后,其底物一側均為還原狀態,其氧一側均為氧化態,這很容易用分光亮度法(雙波長分光亮度計)檢定,重要的呼吸抑制劑有以下幾種。
魚藤酮(rotenone)系從植物中分離到的呼吸抑制劑,專一抑制NADH→CoQ的電子傳遞。
抗霉素A(actinomycinA)由霉菌中分離得到,專一抑制CoQ→Cyt c的電子傳遞。
(二)磷酸化抑制劑 這類抑制劑抑制ATP的合成,抑制了磷酸化也一定會抑制氧化。
寡霉素(oligomycin)可與F0的OSCP結合,阻塞氫離子通道,從而抑制ATP合成。
二環己基碳二亞胺(dicyclohexylcarbodiimide,DCC)可與F0的DCC結合蛋白結合,阻斷H+通道,抑制ATP合成。櫟皮酮(quercetin)直接抑制參與ATP合成的ATP酶。
(三)解偶聯劑(uncoupler) 解偶聯劑使氧化和磷酸化脫偶聯,氧化仍可以進行,而磷酸化不能進行,解偶聯劑作用的本質是增大線粒體內膜對H+的通透性,消除H+的跨膜梯度,因而無ATP生成,解偶聯劑只影響氧化磷酸化而不干擾底物水平磷酸化,解偶聯劑的作用使氧化釋放出來的能量全部以熱的形式散發。動物棕色脂肪組織線粒體中有獨特的解偶聯蛋白,使氧化磷酸化處于解偶聯狀態,這對于維持動物的體溫十分重要。
常用的解偶聯劑有2,4-二硝基酚(dinitrophenol,DNP),羰基-氰-對-三氟甲氧基苯肼(FCCP),雙香豆素(dicoumarin)等,過量的阿斯匹林也使氧化磷酸化部分解偶聯,從而使體溫升高。
過量的甲狀腺素也有解偶聯作用,甲狀腺素誘導細胞膜上Na+-K+-ATP酶的合成,此酶催化ATP分解,釋放的能量將細胞內的Na+泵到細胞外,而K+進入細胞,Na+-K+-ATP酶的轉換率為100個分子ATP/秒,酶分子數增多,單位時間內分解的ATP增多,生成的ADP又可促進磷酸化過程。甲亢病人表現為多食、無力、喜冷怕熱,基礎代謝率(BMR)增高,因此也有人將甲狀腺素看作是調節氧化磷酸化的重要激素。
六、氧化磷酸化的調節
機體的氧化磷酸化主要受細胞對能量需求的調節
(一)ATP/ADP值對氧化磷酸化的直接影響 線粒體內膜中有腺苷酸轉位酶,催化線粒體內ATP與線粒體外ADP的交換,ATP分子解離后帶有4個負電荷,而ADP分子解離后帶有3個負電荷,由于線粒體內膜內外有跨膜電位(△ψ),內膜外側帶正電,內膜內側帶負電,所以ATP出線粒體的速度比進線粒體速度快,而ADP進線粒體速度比出線粒體速度快。Pi進入線粒體也由磷酸轉位酶催化,磷酸轉位酶催化OH與Pi交換,磷酸二羧酸轉位酶催化Pi2-與二羧酸(如蘋果酸)交換。
當線粒體中有充足的氧和底物供應時,氧化磷酸化就會不斷進行,直至ADP+Pi全部合成ATP,此時呼吸降到最低速度,若加入ADP,耗氧量會突然增高,這說明ADP控制著氧化磷酸化的速度,人們將ADP的這種作用稱為呼吸受體控制。
機體消耗能量增多時,ATP分解生成ADP,ATP出線粒體增多,ADP進線粒體增多,線粒體內ATP/ADP值降低,使氧化磷酸化速度加快,ADP+Pi接受能量生成ATP。機體消耗能量少時,線粒體內ATP/ADP值增高,線粒體內ADP濃度減低就會使氧化磷酸化速度減慢。
(二)ATP/ADP值的間接影響 ATP/ADP值增高時,使氧化磷酸化速度減慢,結果NADH氧化速度減慢,NADH濃度增高,從而抑制了丙酮酸脫氫酶系、異檸檬酸脫氫酶、α-酮戊二酸脫氫酶系和檸檬酸合成酶活性,使糖的氧化分解和TCA循環的速度減慢。
(三)ATP/ADP值對關鍵酶的直接影響 ATP/ADP值增高會抑制體內的許多關鍵酶,如變構抑制磷酸果糖激酶、丙酮酸激酶和異檸檬酸脫氫酶,還能抑制丙酮酸脫羧酶、α-酮戊二酸脫氫酶系,通過直接反饋作用抑制糖的分解和TCA循環。
七、高能磷酸化合物的儲存和利用
無論是底物水平磷酸化還是氧化磷酸化,釋放的能量除一部分以熱的形式散失于周圍環境中之外,其余部分多直接生成ATP,以高能磷酸鍵的形式存在。同時,ATP也是生命活動利用能量的主要直接供給形式。
(一)高能化合物
人體存在多種高能化合物,但這些高能化合物的能量并不相同。
體外實驗中,在pH7.0,25℃條件下,每克分子ATP水解生成ADP+Pi時釋放的能量為7.1千卡或30.4千焦耳,在體內,pH7.4,37℃,ATP、ADP+Pi、Mg2+均處于細胞內生理濃度的情況下,每克分子ATP水解生成ADP+Pi時釋放的能量為33.5-50千焦耳或8-12千卡(表6-4)。
表6-4 幾種常見高能化合物水解時釋放的能量
| 化合物 | 千焦耳/克分子 | 千卡/克分子 |
| 磷酸烯醇式丙酮酸 | -62.1 | -14.8 |
| 1,3-二磷酸甘油酸 | -49.5 | -11.8 |
| 磷酸肌酸 | -43.9 | -10.5 |
| 乙酰CoA | -31.4 | -8.2 |
| ATP | -30.4 | -7.3 |
| S-腺苷蛋氨酸 | -29.3 | -7.0 |
| F-6-P | -15.6 | -3.8 |
| 谷氨酰胺 | -14.2 | -3.4 |
| G-6-P | -13.48 | -3.3 |
衛生學規定,中度體力勞動者每日kg體重需供給能量34-40千卡,若一成人重70kg,從事中度體力勞動,則每日應供應含能量2450千卡的食物,其中40%的能量轉變成化學能儲存于ATP分子的高能鍵中,這一部分能量應為2450×0.4=980.0千卡,按每克分子ATP水解生成ADP+Pi釋放7.3千卡能量計算,應當合成:980÷7.3=134.3克分子ATP,ATP的分子量為507.22,所以134.3克分子ATP重達68.12kg,這足以表明ATP在體內的代謝十分旺盛。
ATP在能量代謝中之所以重要,就是因為ATP水解時的標準自由能變化位于多種物質水解時標準自由能變化的中間,它能從具有更高能量的化合物接受高能磷酸鍵,如接受PEP、1,3-二磷酸甘油、磷酸肌酸分子中的~Pi生成ATP,ATP也能將~Pi轉移給水解時標準自由能變化較小的化合物,如轉移給葡萄糖生成G-6-P。
(二)ATP能量的轉移
ATP是細胞內的主要磷酸載體,ATP作為細胞的主要供能物質參與體內的許多代謝反應,還有一些反應需要UTP或CTP作供能物質,如UTP參與糖元合成和糖醛酸代謝,GTP參與糖異生和蛋白質合成,CTP參與磷脂合成過程,核酸合成中需要ATP、CTP、UTP和GTP作原料合成RNA,或以dATP、dCTP、dGTP和dTTP作原料合成DNA。
作為供能物質所需要的UTP、CTP和GTP可經下述反應再生:
UDP+ATP→UTP+ADP
GDP+ATP→GTP+ADP
CDP+ATP→CTP+ADP
dNTP由dNDP的生成過程也需要ATP供能:
dNDP+ATP→dNTP+ADP
(三)磷酸肌酸
ATP是細胞內主要的磷酸載體或能量傳遞體,人體儲存能量的方式不是ATP而是磷酸肌酸。肌酸主要存在于肌肉組織中,骨骼肌中含量多于平滑肌,腦組織中含量也較多,肝、腎等其它組織中含量很少。
磷酸肌酸的生成反應如下:
![]()
肌細胞線粒體內膜和胞液中均有催化該反應的肌酸激酶,它們是同工酶。線粒體內膜的肌酸激酶主要催化正向反應,生成的ADP可促進氧化磷酸化,生成的磷酸肌酸逸出線粒體進入胞液,磷酸肌酸所含的能量不能直接利用;胞液中的肌酸激酶主要催化逆向反應,生成的ATP可補充肌肉收縮時的能量消耗,而肌酸又回到線粒體用于磷酸肌酸的合成,此過程可用圖6-15表示。
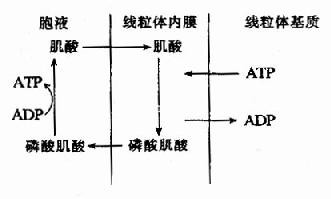
圖6-15 磷酸肌酸的生成與利用
肌肉中磷酸肌酸的濃度為ATP濃度的5倍,可儲存肌肉幾分鐘收縮所急需的化學能,可見肌酸的分布與組織耗能有密切關系。
ATP的生成、儲存和利用可用下圖表示(圖6-16)
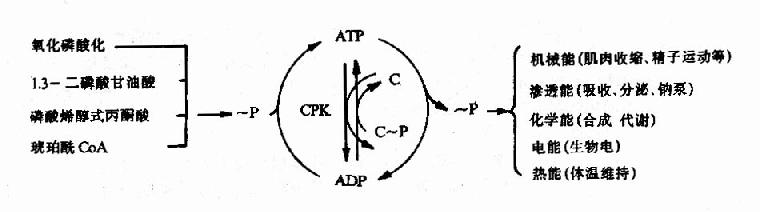
圖6-16 ATP的生成、儲存和利用總結示意圖
CPK:肌酸磷酸激酶
| 關于“生物化學與分子生物學/ATP的生成、儲存和利用”的留言: | |
|
目前暫無留言 | |
| 添加留言 | |